Función del calcio
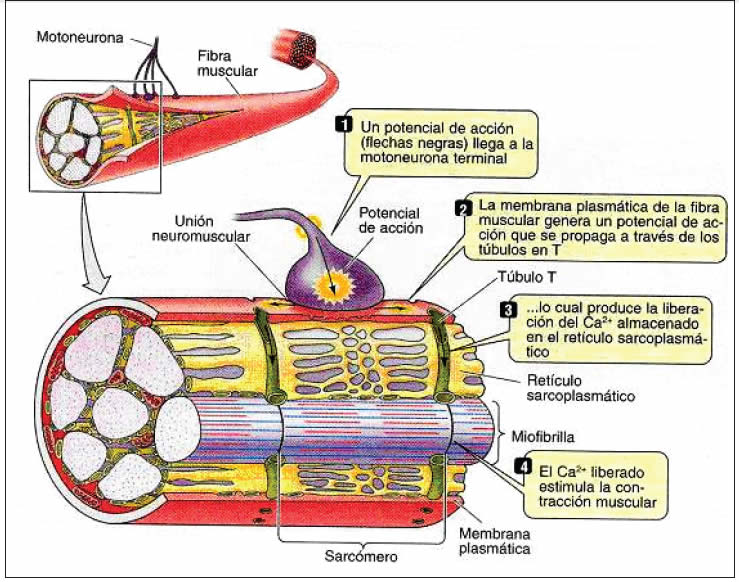
Para iniciar la contracción muscular, los canales de calcio presentes en la membrana del retículo sarcoplásmico se abren porque a través de los túbulos T ingresa el potencial de acción desde el sarcolema (membrana de a fibra muscular), permitiendo que este calcio pase al sarcoplasma y se una a la subunidad C de la troponina (Figura 12 B). La unión provoca un cambio de conformación de la troponina C, que saca a la tropomiosina llevándola hacia la hendidura y descubriendo los sitios de unión entre la actina y la miosina (filamento delgado “on”. La cabeza de la miosina que se había asociado e hidrolizado a una molécula de ATP, aún mantenía los productos ADP y Pi unidos, al asociarse a la actina libera estos productos, lo que genera un “tirón” de la miosina sobre la actina. Los filamentos delgados son llevados hacia el centro d el sarcómero, por lo cual el sarcómero se acorta y se genera tensión activa. Si no siguen otros potenciales de acción, el calcio es bombeado de vuelta hacia sus reservorios, la troponina C recupera su conformación de reposo, la tropomiosina vuelve a cubrir el sitio de unión de actina con miosina. Esta última se “recarga” con ATP y el músculo se relaja (Figura 12). La energía es suministrada por la hidrólisis del ATP. El proceso de hidrólisis del ATP por la ATPasa de la miosina es acelerada por la unión de la cabeza de la miosina a la actina.
El ciclo bioquímico de la contracción muscular se explica en la Figura 13.
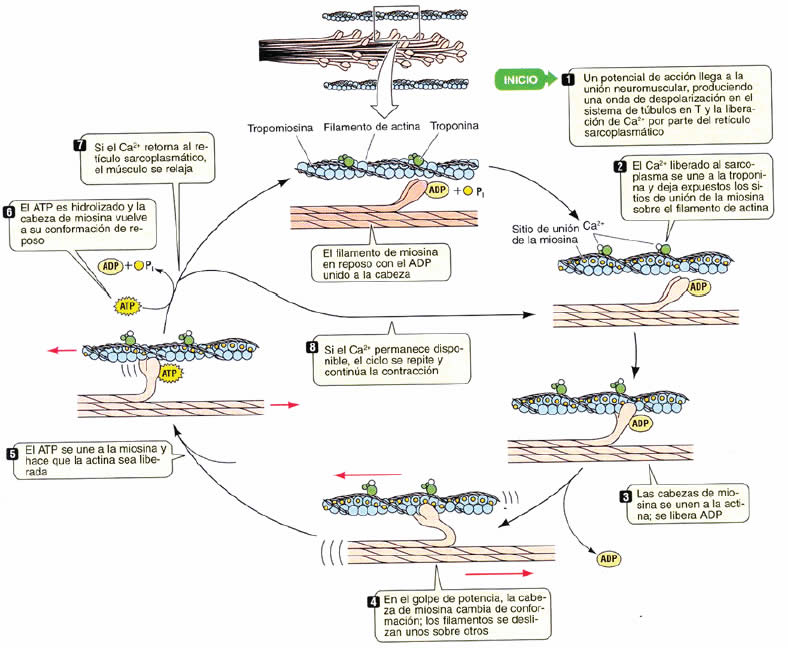
De forma clara el ATP separa la cabeza de miosina del filamento delgado y le da la energía a la contracción.
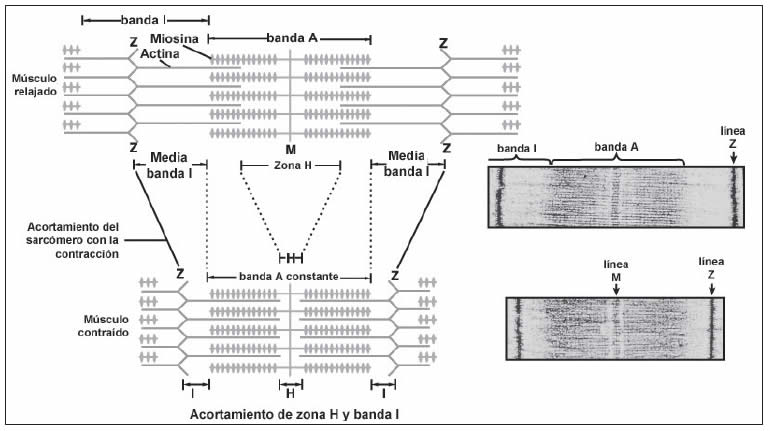
El proceso mediante el cual se realiza el acortamiento de los elementos contráctiles en los músculos implica el deslizamiento de los filamentos delgados sobre los gruesos. El ancho de la banda A permanece constante, en tanto que las líneas Z se juntan cuando el músculo se contrae y se separan cuando se relaja. Cuando el músculo se acorta, los filamentos delgados se aproximan entre sí desde los extremos opuestos del sarcómero; cuando el acortamiento es marcado, estos filamentos se traslapan (Figura 14).
Recordemos que la concentración sarcoplásmica de ion calcio es muy baja en reposo (del orden de 10-8 M). Cuando cada potencial de acción invade el terminal axonal, provoca la liberación de cerca de 300 vesículas ricas en acetilcolina del terminal. Cada vesícula contiene cerca de 10.000 moléculas de este neurotransmisor.
La acetilcolina liberada difunde por el espacio sináptico. Cerca de un tercio de ella es hidrolizada por la acetilcolinesterasa neural antes de alcanzar los receptores musculares. Cada pareja de moléculas de acetilcolina que alcanza un receptor provoca la entrada de cerca de 50.000 iones sodio, porque se abren los canales para este ión, provocando la despolarización del sarcolema.
Los potenciales de acción musculares se propagan por el sarcolema y penetran al interior del músculo por el sistema tubular transversal, llamado sistema T y provocan, por un mecanismo aún no del todo claro, la liberación del calcio contenido en las cisternas terminales (Figura 15).
En el corazón el proceso es muy parecido, excepto que el potencial no se genera por la activación de las motoneuronas, sino por la acción de las células del marcapaso. Otra característica muy importante del músculo cardíaco es que posee un período refractario muy largo y por esta razón no se tetaniza (es decir, no entra en contracción mantenida, lo que impediría la circulación de la sangre). En el músculo estríado, los filamentos delgados están insertos en la línea Z, mientras en el músculo liso están en los llamados cuerpos densos, que se encuentran en general adosados a la membrana celular. No existe aquí la fina estructuración del sarcómero, donde cada filamento grueso está rodeado por seis filamentos delgados en una configuración hexagonal. De este modo, la contracción del músculo liso es en todas las direcciones del espacio y no en forma lineal como ocurre en el músculo estríado. Por otra parte, no están presentes en el músculo liso las proteínas reguladoras (troponinas, tropomiosina) y es más abundante la actina que la miosina, lo que asemeja mucho la contracción del músculo liso a los movimientos citoesqueléticos, también dependientes de actina.