Bases iónicas del potencial de acción
Bases iónicas del potencial de acción
Si se aplica un estímulo de cierta magnitud en la membrana de una célula excitable, se produce un ligero incremento en la permeabilidad de los iones sodio en esa región disminuyendo levemente la diferencia de potencial de acuerdo a la intensidad del estímulo. Un estímulo umbral es aquel que posee la intensidad suficiente para producir una disminución en el voltaje, aproximándose a los -55 mV que se denomina nivel de descarga, voltaje en el cual se abren los canales de sodio permitiendo la entrada masiva del ión (canal tipo compuerta de voltaje), de manera que el voltaje del medio intracelular se va acercando a cero o sea se provoca una despolarización. La masiva entrada del Na+ hace que el lado interno de la membrana plasmática quede positiva alcanzando +35 mV: potencial de espiga. En ese instante los canales de sodio se cierran rápidamente (terminando la entrada masiva de sodio) y se abren totalmente los canales de potasio (que ya se habían comenzado a abrir lentamente) determinando la salida de este ión, lo que vuelve a hacer negativo el lado intracelular de la membrana: repolarización, luego de una ligera hiperpolarización (debido a la salida de potasio en ausencia de la entrada de sodio),la acción de la bomba Na+-K+ ATP asa permite alcanzar nuevamente el potencial de reposo.
Regionalmente, después de un potencial de acción, la posición relativa de los iones sodio y potasio está invertida, la acción de la bomba sodio-potasio restituye las posiciones originales. Mientras se conduce un potencial de acción obviamente no puede conducirse otro, y un estímulo en este período no origina un nuevo potencial de acción (período refractario absoluto).
Aunque la mayoría de las células tienen potencial de reposo, sólo dos de ellas, la neurona y la célula muscular, pueden experimentar fácilmente impulsos electroquímicos, también llamados potenciales propagados o potenciales de acción, constituyendo los tejidos excitables. Esto se debe a que estos tipos celulares (además de la citada bomba) poseen en su membrana canales iónicos (proteínas integrales de membrana específicas), unos para el sodio y otros para el potasio. La apertura de estos canales, como respuesta al estímulo, permite el libre tránsito de los iones de acuerdo a sus gradientes.
Durante la propagación el impulso nervioso no pierde intensidad desarrollando siempre el mismo potencial de acción. En vivo los potenciales de acción se propagan unidireccionalmente en el axón (desde el soma hacia el telodendrón), sin embargo, si experimentalmente se aplica el estímulo sobre algún punto del axón, este se propaga en ambas direcciones.
Es importante observar que si el estímulo inicial, no hubiese tenido la magnitud suficiente para producir una disminución en el potencial de membrana cercana a -55mV, los canales de sodio no se hubiesen abierto completamente y el trabajo de la bomba sodio-potasio restablecería el potencial inicial, en esta situación se estaría frente a un estímulo subumbral. Por otra parte, si el estímulo inicial hubiese sido de un registro superior al necesario, Estímulo Supraumbral, la magnitud de descarga habría sido la misma que con un estímulo umbral, esto se denomina Ley del Todo o Nada.
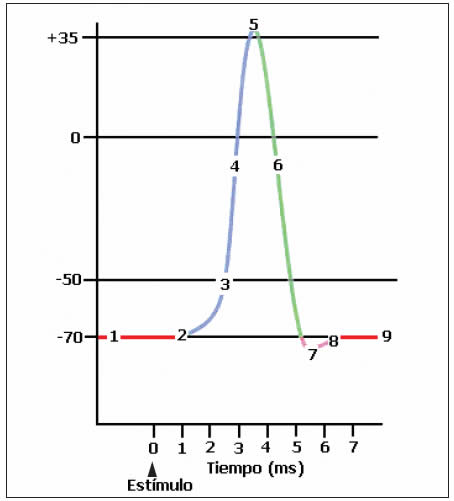
1. Potencial de reposo.
2. Estímulo despolarizante.
3. La membrana se despolariza al llegar al umbral. Se abren los canales de Na+ voltaje-dependientes y el Na+ ingresa. Los canales de potasio comienzan a abrirse lentamente.
4. La rápida entrada de Na+ despolariza la célula.
5. Se cierran canales de Na+ y se abren lentamente los de K+.
6. El K+ sale hacia el fluido extracelular.
7. Los últimos canales de K+ se abren y el ion sale, provocando la hiperpolarización.
8. Cierre de los canales de K+ voltaje dependientes y algunos iones de K+ ingresan a la célula a través de canales. Acción de la bomba Na+ /K+.
9. La célula retorna a su estado de reposo, recuperando su potencial de reposo.
La velocidad de conducción de una fibra nerviosa depende principalmente de dos aspectos:
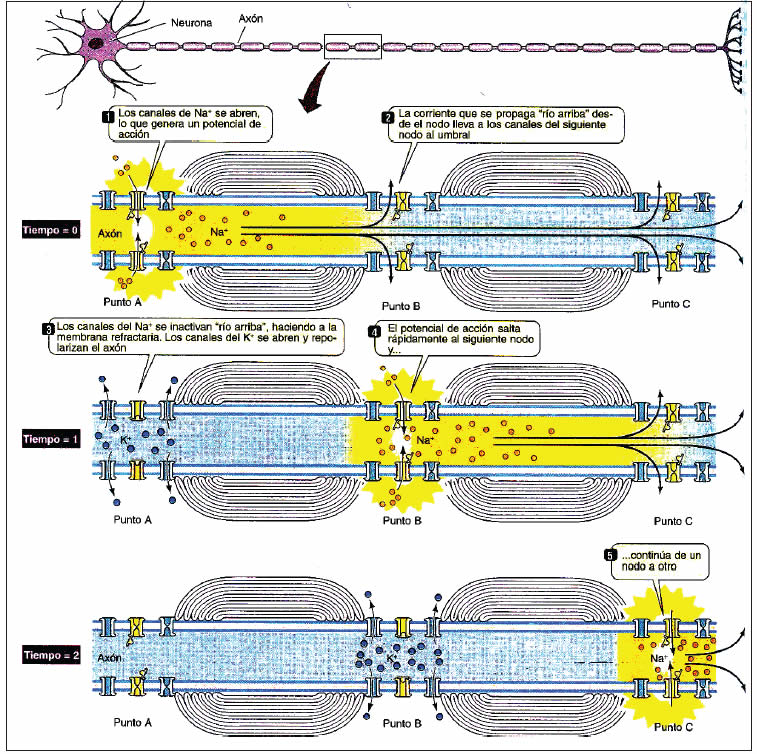
a) Desarrollo de una vaina de mielina: que deja sólo algunas zonas del axolema (membrana citoplasmática de la neurona) descubiertas. En este caso la zona a repolarizar es muy pequeña, y se gana en velocidad de conducción utilizando la llamada “conducción saltatoria”. En la fibra mielínica los canales para iones sensibles a potencial se ubican en la zona amielinizada, nodos de Ranvier. La despolarización de un nodo provoca una “corriente en remolino” que despolariza al nodo contiguo. Así, el potencial de acción cursa por la fibra a una gran velocidad. Una ventaja adicional de la conducción saltatoria es la menor entrada y salida neta de iones sodio y potasio respectivamente, ahorrando energía en la restitución de los iones a sus compartimientos y consiguiendo además períodos refractarios más cortos (Figura 7).
b) Diámetro: Un modo de aumentar la velocidad de conducción es mediante el aumento del diámetro en los axones amielínicos, ya que incrementa la superficie de intercambio iónico.
Toxinas que envenenan los canales iónicos
Algunos organismos han desarrollado toxinas específicas de los canales como mecanismos de autodefensa o para capturar presas. Una variada colección de toxinas naturales tiene por blancos selectivos los canales iónicos de las neuronas y otras células. Estas toxinas son útiles no sólo para la supervivencia sino también para estudiar la función de los canales iónicos celulares. La toxina de canal mejor conocida es la tetradotoxina, producida por el pez globo y algunos otros animales. La tetradotoxina provoca una obstrucción potente y específica de los canales del Na+ responsables de la generación del potencial de acción, paralizando así a los animales que lamentablemente la ingieren. La saxitoxina, homólogo químico de la tetradotoxina, tiene una acción similar sobre los canales del Na+. Los efectos potencialmente letales de la ingestión de mariscos que han ingerido dinoflagelados de la “marea roja” se deben a las acciones neuronales potentes de la saxitoxina.
En cambio, la batrachotoxina, alcaloide esteroideo producido por una rana sudamericana, produce una despolarización persistente, lo que induce una parálisis en la conducción axonal por aumento selectivo de la permeabilidad de la membrana al sodio.
Algunas plantas producen toxinas similares, incluidas la aconitina, de los botones de oro; la veratridina,de las lilas, y algunas toxinas insecticidas producidas por plantas tales como los crisantemos y los rododendros.
Los canales del K+ también han sido blancos de los organismos productores de toxinas. Las toxinas peptídicas que afectan los canales del K+ incluyen la dendrotoxina de las avispas; la apamina de las abejas; y la caribdotoxina, que producen los escorpiones. Todas estas toxinas bloquean los canales del K+ como acción primaria; no se conoce ninguna toxina que afecte la activación o la inactivación de estos canales, aunque puede ser que estos agentes existan y que simplemente aún no hayan sido descubiertos.